Behavior Genetics in Context of Russian Psychology
Malykh, S. B.  Psychological Institute of the Russian Academy of Education, Moscow, Russia Lomonosov Moscow State University, Moscow, Russia
Psychological Institute of the Russian Academy of Education, Moscow, Russia Lomonosov Moscow State University, Moscow, Russia
Abstract
The article is devoted to individual differences in psychological characteristics, the influence of genotype and environment on individual differences in evoked potential of brain related to the execution of motor action, the role of genetic and environmental factors in individual characteristics of the parameters of sensorimotor activity, genetic and environmental factors in development of psychological traits.
Themes: Behavioral genetics
PDF: http://psychologyinrussia.com/volumes/pdf/2011/07_2011_malykh.pdf
Pages: 108-128
DOI: 10.11621/pir.2011.0007
Keywords: individual diff erences, genotype, environmental factors, development.
Introduction
A study of the nature of individual differences in psychological characteristics has almost a century of history and an immense empirical material has been presently collected up, pointing the influence of hereditary factors in the emergence of individual differences in intellect, academic achievement, particular cognitive abilities, cognitive styles, temperament, personality traits, psycho-physiological characteristics, movement and so on. Although these data is actively used to explain and understand psychological phenomena, there are very few attempts to build the logic of genetic research within a framework of general psychological concepts and searchings of psychological principles explaining the data obtained, because the main aim of these works are proper genetic goals.
One of the first attempts to analyze the psychological aspects in genetic research was undertaken back in the 30-s by A.R. Luria. A.R. Luria’s experimental study (1935, 1956, 1962, 2002) showed that the influence of genetic factors at different ontogenetic stages of a particular psychic function development vary. Based on a conception of psychological development of L.S. Vygotsky he assumed that this is related to the change in internal mechanisms of execution of this psychological function, to the qualitative rebuilding of psychological activity of a child, forming of complex functional systems in the process of education. This rebuilding, as A.R. Luria considered, consisted in that elemental, immediate forms of activity are replaced by complexly organized functional structures, formed on a basis of a child’s communication with an adult within the process of education. Owing to the acquisition of language the complex forms of psychological activity acquire new qualities and the mechanisms of their execution are changed and are becoming different from those of elemental processes. Therefore it can be assumed that “the nature of every function (in other words – her relation to the genotype) is changed too in a process of human psychological development as well as its structure...” (Luria, 1962, p. 16).
No wonder this approach to the interpretation of genetic data has been used in developmental research, because it is exactly this area where the interests of psychology, behavioural genetics and biology of development maximally intersect.
One has to note that current psychological approaches (P. Baltes, J. Nesselroade, W. Schaie, R. Lerner and others) in developmental research provide theoretical basis for psychological interpretation of data gathered in behavioural genetics. According to the “systemic” approach different “developmental sources” are parts of an integrated system of higher order and they influence each other. From R. Lerner’s (Lerner & Walls, 1999) point of view, the most promising in current developmental psychology is a study of how the functioning of structures and structuring of functions are changed with time.
In contemporary theories of personality development personality is not “biologised”, “psychologised” or “sociologised”, but is regarded from a systemic point of view. Atomistic and mechanistic views are replaced by models underlying dynamic synthesis of multiple levels of analysis, as well as activity nature of human development and systemic nature of development in general.
Development is a quality, an attribute of systemic changes at different interdependent levels of organization. From R. Lerner’s point of view, the basis of changes and limits of development is laid down in relationships among multitude of organizational levels, composing human life. These levels include from “inner biological through individual psychological and proximate social relationships level (including child-parent dyads, peer groups and nuclear families) to socio-cultural (including those key organizations as educational system, social organizations, government and economic systems) and parameters of environment mastered and transformed by a human” (Lerner & Walls, 1999).
None of the organizational levels functions on its own. The functioning specific of every level is conditional on its structural integration in a system of all levels. History – the change with time – never ends and it can be contemplated as a particular level of organization, amalgamated with other levels. It implies that everything changes: the variables’ estimates, structures and functions.
Developmental biology data suggest that the idea of genetic directive is unlikely even for morphogenesis, because the exact cell localization can’t be coded in gene (Edelman, 1992). Genes control developmental processes indirectly, determining which proteins regulating morphogenesis will be produced, but “the real microscopic destiny of a cell is determined by epigenetic events, which depend on developmental history unique for every individual embryonic cell” (Ibid., р. 62). Thus, Edelman introduces a concept of historical randomness as a part of a theory of embryonic development.
Development, according to current views, is a systemic dynamic process, in which genes might play a role of triggers in processes of cell differentiations, guiding cell development in one or another way, but by that there are many nongenetic factors (cell environment, different signals from other cell systems and outside environment), which modify development (Michel & Moore, 1995).
Evidently psychological trait (phenotype), which has a complex internal structure, is qualitatively different from the traits traditionally studied in genetics; understanding of the nature of psychological phenomena is impossible without analysing of psychological “context”, in which the studied trait is implemented.
According to I.V. Ravich-Scherbo “…psychological traits as objects of genetic analysis have particular qualities, which one can’t not to take into account”, because “events, not structures” are used as phenotypes (Ravich-Scherbo, 1988).
The data of genetic analysis of psycho-physiologic traits enable to trackle a very , as we think, interesting logic – the increase in heritability estimates of some measures in experimental situations requiring a higher degree of a subject’s “involvement” (Boomsma et al., 1990). In most cases it is probably not possible to talk about the heritability of a physiological measure alone, outside the context, in which it is included in a study. The researches using this paradigm are not many, but their strengths are obvious.
Literature data analysis allowed singling out a particular logic of expression of genetic and environmental influences in individual differences of psychological and psycho-physiological traits depending on a “context”, in which the trait is implemented. The data of the whole range of studies suggest that the structure of phenotypic variance of psychological and psycho-physiological traits depends on their psychological structure, on a “context”, in which a trait is implemented, on its place, meaning, functional role in the whole system a trait is included at a moment of investigation (Panteleyeva, 1977; Kochubei, 1983, 1988; Zyryanova, 1992; Malykh, 1986, 1988, 1993, 2000; Malykh et al., 1986, 1993; Maryutina, 1978, 1994, 1988; Talyzina et al., 1991; Egorova, 2000, Boomsma et al., 1992, 1998; Fulker et al., 1993; van Baal, 1997 etc). The results of these studies demonstrate that the same behavioral trait can have qualitatively different execution mechanisms, by that in every particular case different links of functional system can be recruited.
These data determined the logic of experimental studies – the use of such experimental schemes that provide variations of psychological content of activity while measuring the same psychological trait – analysis of age dynamics of genetic determinants of psychological traits in the course of individual development and brain mechanisms of psychological human functioning. In the following account we stick to this logic.
The influence of genotype and environment on individual differences in evoked potential of brain related to the execution of motor action
To estimate the effect, which a movement’s localization in a structure of a whole motor action has on a proportion of genetic and environmental factors within individual variations of central mechanisms of construction and execution of motor action, special experimental schemes have been built. The same movement with the same biomechanical structure (pushing a button), but taking different place in a functional system of a whole motor action has been investigated (Malykh, 1986; Malykh et al., 1986).
A method of registration of a complex of brain potentials related to the movement (MRBP) has been used for analysis of central mechanisms related to the preparation and execution of motor action. The processes of preparation, execution and control of a movement are reflected in a complex of potentials, preceding, execution and estimation of a movement.
25 monozygotic (MZ) twins and same-sex dizygotic (DZ) twins aged 18-30 years took part in the study. Amplitude and time characteristics of c MRBP components have been analyzed in three experimental situations: by simple voluntary movements (1), by pushing a button in a situation of a prognosis of two signals of an equal probability (2) and in a situation of a true prognosis (3).
The results of genetic analysis revealed that genetic influences are more oft en present in a situation of a prognosis of two signals of an equal probability (55,9% of MRBP analyzed parameters are described by a simple genetic model), while simple voluntary movements are considerably more rare (21,4%). The situation of a true prognosis takes an intermediate place (39,7%).
Therefore despite the fact that in all experimental situations a biomechanically identical movement has been used the dynamics of genetic control of neuro-physiological correlates of movement organization depending on a type of experimental situation, on a different psychological context of a movement execution. In a first experimental situation pushing a button is an independent act of voluntary human activity appearing as a conscious goal. A movement in this case is an action, because an action as an activity entity is a process subservient to the understanding of a result, which should be achieved, in other words a process, subservient to a conscious goal. In a prognosis situation the same movement becomes the only one of the conditions of a new action determined by a new goal (to predict the location a next signal), in other words a way of executing of a new action, according to A.N. Leontjev, an operation, its executing, and it stops to be executed as a goal-oriented process: its goal is no more distinguished. Thus a movement, which has been a goal of an action, becomes the means of executing another action. By that despite preservation of a biomechanical schema, the organization of an action in a central nervous system will take place, according to N.A. Bernstein, at different levels of movement construction. Therefore, according to the change of a role of movement in a structure of the whole action, the mechanisms of its regulation are changed, that leads to the changes in a role of genetic and environmental factors in a variation of MRBP.
Age dynamics of MRBP genetic control in a course of development
Genetic analysis of age dynamics of MRBP at different developmental stages can serve as a useful paradigm for an analysis of dependency of hereditary determination of BPRM on psychological structure of a studied trait, because the execution mechanisms of psychological function change in the course of ontogenesis. With this goal in mind we conducted a series of studies (Kuznecova, 1992; Malykh & Kuznecova, 1992; Orekhova, 1996; Malykh, 1986). Unfortunately, ontogenetic studies of MRBP are infrequent and fragmented, yet existing research suggest change of MRBP parameters in the course of ontogenesis.
29 pairs of MZ and 32 pairs of DZ twins aged 6-8 years, 23 pairs of MZ and 16 pairs of DZ twins aged 8-11 years, 22 pairs of MZ and 17 pairs of DZ twins aged 11-13 years and 25 pairs of MZ and 25 pairs of DZ twins aged 18-30 years participated in this experimental series. BPRM has been registered in a situation of a simple voluntary movement.
In fact, age analysis revealed that MRBP is changing considerably from the age of 6 to the age of 13, these changes are related to the specifics of brain organization of motor action at different age periods (Malykh et al., 1992; Malykh & Orekhova, 1995). By the age of 13 MRBP doesn’t commonly reach definitive value. The age dynamics of MRBP observed in a diapason of 6-13 years as well as the differences between children and adults reflect maturation of brain mechanisms of motor control and probably are determined by a number of psychological factors (attention level, movement automation, strategy of a motor action and so on) as well.
Comparison of data on the influence of genetic and environmental factors on variability of MRBP amplitude characteristics of children aged 6-13 and adults showed that with age there is an increase in a number of parameters, whose variability is better described by a simple genetic model: in 6-8 years this number is 25%, in 8-11 years – 31,25%, in 11-13 years – 43,75% and in 18-30 years – 62,5% of parameters (Malykh, 2000).
Overall comparison of data obtained on child and adult populations suggest that proportion of genetic and environmental influences in individual characteristics MRBP changes with age; that is reflecting a change of brain mechanisms of motor action organization, which takes place in a course of ontogenesis.
The role of genetic and environmental factors in individual characteristics of the parameters of sensorimotor activity
A special experimental study has been undertaken to cover the dynamics of gene-environment effects in individual differences of the parameters of sensorimotor activity when varying separate links of the contour of self-regulation (Ivanchenko & Malykh, 1994; Ivanchenko, 1995).
The problem of self-regulation has a manifold character and touches all levels of organization, starting from neurophysiological to the personality regulation of activity. The works of N.A. Bernstein, P.K. Anokhin, and K. Pribram became fundamental for our understanding of goaloriented and purposeful behaviour of living systems; owing to these works it became possible to apply physiological and cybernetic models to the analysis of psychological mechanisms of self-regulation.
Psychological mechanisms of conscious regulation of activity were extensively investigated in a set of studies supervised of O.A. Konopkin (1962-1980). Using such measures as sensory sensitivity, reaction time (RT) as examples, a fact that the dependence of sensorimotor activity on a complex of objective conditions, in which it is executed, is mediated by a process of goal-oriented regulation has been theoretically formulated and experimentally proved (Konopkin О.А., Morosanova V.I., Osnicky А.К., Stepansky V.I.; Zhuikov Yu.S., Sipachov N.О.; Kondratieva I.I., Mislavsky Yu.I.). Specifically the dependence of RT on stimulus intensity (physiological “law of power”) is expressed differently in situations of random and ordered signals’ presentation and is influenced by the probability of emergence of particular signal, duration of inter-stimuli interval, interval between a warning and triggering signals (Boyko V.I.). But these objective laws also change under the influence of such factors as a goal of an action, subjective fullness of a model of activity conditions, presence or absence of a success criteria and so on (Kondratieva, 1973; Olshannikova, 1964).
It can be assumed that to describe a complicated structure of genotype- environmental balance in determination of a concrete trait the model can be successfully employed, which reflects a holistic picture of activity, in which this trait is included. A functional structure of a system of conscious regulation of activity can serve as a sort of the model (Konopkin, 1980).
40 pairs of MZ and 40 pairs of same-sex DZ twins aged 9-14 years took part in this experimental series. The essence of this experiment consisted in a variation of the conditions of presentation to the participants of auditory stimuli of different intensity, reaction to which was an elemental movement act (pushing a button). Every subject’s time of sensorimotor reaction to the sound was measured in 6 experimental situations, in which the order of auditory signals, instruction and a way of response action were varied:
In the 1st and 2nd series stimuli were presented randomly, but with a different instruction “React when you hear a signal” (1) and “as fast as possible” (2).
In the 3rd series stimuli were presented in a descending order by five volume levels, from a maximal to a minimal, and the instruction “as fast as possible” was kept and in addition the sound of 40 db was singled out as subjectively significant. In the 4th series a subject received information about the current results. In series 3 and 4 the role of the links of subjective model of conditions was evaluated.
In series 5 and 6 the role of the link of programming of execution actions was tested. In these series the reaction of choice from three alternatives was used. In the 5th series the order of signals was descending, while in the 6th it was random.
In all tasks the auditory signals of five intensity levels (40, 60, 80, 100, 115 db) were presented to the subjects. Stimulus raw consisted of 80 signals; a sound of every intensity level appeared 16 times. Also an “empty” signal, a warning flash without a following auditory signal appeared 16 times. A response reaction was pushing a button with a right hand.
Genetic analysis of data (mean RT and coefficient b) was conducted separately for every age group: 9-11 years of age (21 pairs of MZ and 22 pairs of DZ) and 12-14 years of age (19 pairs of MZ and 18 pairs of DZ).
In a younger age group additive genetic effects dominated in tasks 2, 4, 6, and the number of measures, which variability is described by a single genetic model, increased according to the increase in task’s complexity. Contrary to that in “background” tasks 1, 3, 5 predominantly environmental influences are detected; the largest part of these influences is explained by non-shared environmental effects.
In the older age group the proportion of genetic and environmental variance components is somewhat different. The role of shared environment grows in all tasks, except 1 and 5 series, where the variance of RT is explained by additive genetic effects.
Therefore, in a transition from a young age group to the old group we observe a change of genetic-environmental proportion in the variability of RT in all experimental conditions, which reflect the transition to the new level of activity regulation.
The data obtained suggest that the dependence of genetic and environmental factors impact phenotypic variability of RT and coefficient b on experimental conditions. In other words, phenotypically identical measure, for example RT, in different tasks is included in different functional systems. Analysis of two age groups showed the presence of particular age dynamics of genetic control of studies measures, which probably reflects qualitative changes happening in psychomotor sphere in these ages.
Therefore, the data obtained suggest that phenotypically identical trait, meaning a trait possessing the same external expression, might significantly differ in its psychological structure and accordingly in the proportion of the impact of genetic and environmental determinants in its variability.
A study of a role of heredity and environment in individual differences of psychological traits
The data suggesting that the proportion of genetic variance in variability of a psychological trait is closely related to the content and psychological structure of the activity, which performance results are used to measure the studied trait, create particular interest in a study of the operational side of activity. If the subject’s activity during the problem solving stays the same externally, but psychological content of the activity changes, it can express itself in the changes of the architecture of hereditary and environmental proportions. Particularly, the content of psychological trait might change due to the learning process.
To test this assumption we used the data from a study of successfulness of prognosis in a situation before and after the learning (Malykh, Egorova, & P’yankova, 1993a; Malykh, Egorova, & P’yankova, 1993b). Every child had to solve six problems of three complexity levels (36 pairs of MZ and 33 pairs of DZ twins aged 6-7 years participated in a study). Problems of the same complexity level have been constructed according to the same principle, but differ in stimulus material. A child has been presented with a strictly determined stimulus sequences, repeating in a particular order. A subject had to forecast the appearance of every subsequent stimulus. Successful prognosis is possible, when understanding of a rule of consequence construction is achieved. After every problem’s solving the principle of construction of this stimulus consequence has been demonstrated to subjects and learning of the ways of testing of the put forward hypotheses-forecasts was conducted. The transition to the next problem was happening after the complete understanding of the preceding problem. When solving problems in a “before learning” situation a subject acted not knowing yet how to do it. When solving problems in an “after learning” situation it has been enough to apply the already known principle to other stimulus material.
Success of prognostic activity (a quantity of correct forecasts) before and aft er education has been recorded. By the way of influencing the mode of child’s activity learning impacted the successfulness of prognosis.
The analysis of these experimental data allowed estimating the dependence of structure of phenotypic variance of prognosis success measures from mechanisms of its execution. To analyze a structure of phenotypic variance of experimental measures in a situation before and after learning a model-fitting methods have been used.
Model-fitting results revealed that in a “before learning” situation a simple genetic model (AE) is the best fitting one (χ2=0,882 0,5<p<0,7), in an “after learning” situation favours environmental (CE) model (χ2=0,297 0,8<p<0,9). Variance of prognosis success “before learning” is explained by additive genetic effects (52,6%) and non-shared environment effects (47,4%), decreasing the degree of relatives’ similarity. For measures registered in an “after learning” situation, variance is explained by shared environment effects (52,2%), increasing the degree of relatives’ similarity and non-shared environment (47,8%).
Thus, the results of the research suggest that etiology of phenotypically the same trait – prognosis success – is different in two experimental situations. In a “before learning” situation genetic influences are clearly expressed, while in an “after learning” situation the acquisition of an effective behavioral strategy leads to the increase in shared environmental factors. In a “before learning” situation a child is acting in a new situation unknown to him, when the methods of success achievement are unknown. You need to find them by yourself, by the way of constructing a cognitive schema adequate for a problem. In a second situation a child can use cognitive schemas already constructed in a process of preliminary goal oriented learning.
Therefore, in the two described experimental situations a child adopts two different strategies of problem solving, choosing qualitatively different modes to achieve a goal. One and the same activity – prognosis of a subsequent event – is executed by using different psychological mechanisms; that leads to the changes in heritability estimates for those psychological traits, which are measured according to the results of this activity execution.
Genetic and environmental factors in development of psychological traits
The data of a recently conducted longitudinal twin study have been used to analyze the dynamic of genetic control in development of psycho logical traits. This study has been conducted by the laboratory of genetic psychophysiology for more than 15 years (from 6 to 21 years of age).
One kind of a simplex-models proposed by L. Guttman has been used to estimate the role of genetic factors in a process of development (Guttman, 1954). The model is presented as a pathway one in a Fig. 1.
The following sources of individual differences were considered: genetic effects (G), common or shared environment (С) and non-shared environment (Е). In 6,5; 7,5; 13,5 and 16,5 years of age two sets of latent factors, which included three effects, were introduced. Covariance of IQ measures in 6,5; 7,5; 13,5 and 16,5 years of age was modeled with latent factors G, C and E, influencing IQ in 6,5, as well as in 7,5, 13,5 and 16,5 years of age. A second set of latent factors described age-specific genetic and environmental effects in a corresponding age.
Four waves of longitudinal study data have been used for the analysis – in 6,5 years of age (47 MZ and 47 DZ pairs), in 7,5 years of age (42 MZ and 35 DZ pairs), in 13,5 years of age (90 MZ and 66 DZ pairs) and in 16,5 years of age (75 MZ and 55 DZ pairs). A level of psychometric intellect – a traditional for behavioral genetics instrument – which was measured by a children version of Wechsler’s test (WISC) – was used.
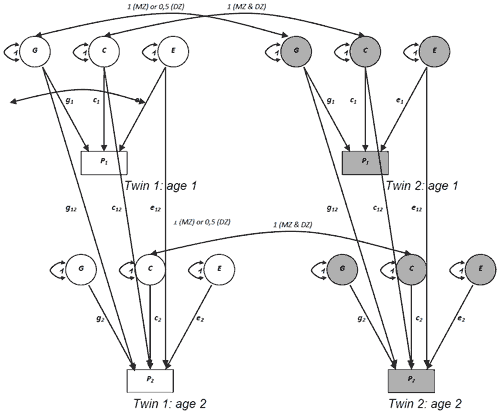
Figure 1. Path diagram representing the simplex model for IQ longitudinal study
The results of genetic analysis of psychometric intellect showed a sufficiently substantial influence of genetic factors on variability of psychometric intellect. Moreover in the course of development the influence of these factors rises, while the influence of shared environment decreases (table 1).
The tracing of children’s cognitive development from the age of 6 to 16 confirmed the presence of a common genetic factor in individual differences of IQ, appearing at the age of 6 and preserving its action until the age of 16. At the age of 6 the influence of genetic factors was pronounced (45%), whereas at the age of 7 the influence of genetic factors sharply decreased to 20%, while the influence of shared environment, on the contrary, increased. In the course of further development the influence of genetic factors on IQ variance increases (from 42% at the age of 13,5 to 57% at the age of 16,5), while the shared environment effects decrease (37% and 30%, accordingly).
Table 1. Genetic (G), shared environment (С) and non-shared environment (E) variance at the age of 6,5; 7,5; 13,5 and 16,5; genetic (G) and shared environment (C) covariance between different ages, the influence of new genetic and shared environment factors
|
Phenotypic variance and covariance |
Components of variance and covariance |
New influences | |||
|
G |
C |
E |
G |
C | |
|
Variance at 6,5 years |
45% |
40% |
15% |
– |
– |
|
Variance at 7,5 years |
20% |
69% |
12% |
– |
66% |
|
Variance at 13,5 years |
42% |
37% |
21% |
– |
– |
|
Variance at 16,5 years |
57% |
30% |
13% |
33% |
– |
|
Covariance between 6 and 7 years of age |
49% |
51% |
0% |
– |
– |
|
Covariance between 6 and 16 years of age |
100% |
0% |
0% |
– |
– |
|
Covariance between 14 and 16 years of age |
64% |
32% |
4% |
– |
– |
The fact of abrupt decrease in heritability estimates at the age of 7 is remarkable considering the general trend of steady increasing genetic influences.
In a number of studies a similar trend of decreasing genetic influences around the age of 7 was revealed. For example, the analysis of Louisville Twin Study (LTS) data showed that heritability monotonously increases from the age of 1 to the age of 6 (at the age of 1 – 10%, at the age of 2 – 16%, at the age of 3 – 18%, at the age of 4 – 24%, at the age of 5 – 38%, at the age of 6 – 54%). At the age of 7 genetic influences decrease to 50%, at the age of 8 and 9 – to 34% and 36% correspondingly, at the age of 14 genetic influences again increase to 68% (Wilson, 1983). In the analysis of compiled data from Colorado Adoption Project (CAP), Twin Infants Project (TIP) and McArthur Longitudinal Twin Study (MALTS), conducted by L. Cardon and his colleagues (1992), heritability estimate was .55 at the age of 1, .68 at the age of 2, .59 at the age of 3, .53 at the age of 4 and .52 at the age of 7. The tendency to the decrease in genetic influences at the age of 7 came forward more clearly in another analysis of compiled CAP, TIP and MALTS data (Fulker et al., 1993). In this analysis the heritability estimate in a best-fitting model increased from .53 at the age of 1 to .57 at the age of 4, and then it fell to .43 at the age of 7, and then increased again to .74 at the age of 9 (Patrick, 2000).
We found that in a period between 6 and 7 years of age the proportion of genetic influences decreases, while that of shared environment increases. We think that it could be due to the beginning of schooling putting requirements on cognitive abilities of children. Results obtained on American sample also support this hypothesis, because children, who participated in these studies, started school at the age of 7 too. Thus schooling leads to the changes in the proportion of genetic and environmental determinants of IQ variability: the beginning of schooling leads to increase in shared environment influences and decrease in the role of heritability. The results obtained in a Dutch sample (Boomsma & Van Baal, 1998) support this reasoning. Heritability estimates obtained in Dutch population twins aged 5 were low (27%) comparing to those obtained in the majority of other studies (Cardon et al., 1992; Fulker et al., 1993; Wilson, 1983) and approached estimates obtained in our sample when twins where 7 years of age (19%). Low heritability estimates obtained in a Dutch study of 5 year-old twins also have relationship to the timing of a start of schooling, because children in Nederland start school at the age of 5. Results obtained at the age of 7 showed the increase in genetic effects, which achieved estimates obtained in the American sample (50%).
Probably the abrupt increase in the range of environmental factors directly related to cognitive development leads to decrease in heritability estimates and increase in shared environment effects, this phenomenon is observed only in the first year of school. Interestingly enough similar changes analyzed in preceding experimental series are noticed under individualized education as well – the acquirement of an effective behavioral strategy leads to increase of shared environmental factors.
Specific analysis of environmental influence (the estimation of the flows’ intensities) suggests that before schooling environment promotes convergence of IQ, rather than its divergence. At school situation changes: environment promotes divergence rather than convergence. It is notable that after the start of schooling the density of divergence increases while the density of convergence diminishes (Kuravsky & Malykh, 2000; Kuravsky & Malykh, 2001). Therefore these results suggest the increasing role of non-shared environment in the emergence of variability in psychometric intellect.
Longitudinal design of behavioral genetic studies allowed us to estimate not only the degree of stability and change in IQ development, but also the degree, in which the observed stability and change is explained by genetic differences between individuals or differences in environmental conditions, in which children are brought up. The analysis showed that the impact of genetic and shared environmental factors in stability of development is significant, and each of them explains significant fraction of covariance of cognitive measures. The decomposition of IQ covariance between ages 6 and 7, 13,5 and 16,5 in genetic and environmental components showed that the largest part (49% and 64% accordingly) of covariance is explained by the stability of genetic factors.
The majority of studies suggest the stability of genetic influences on general cognitive characteristics (for example, Boomsma & Van Baal, 1998; Cherny et al., 1998; Fulker et al., 1993). Although the absence of new genetic variance at the age of 7 comply with the results from Louisville twin study (Eaves et al., 1986) and the research of D. Boomsma and C. Van Baal (1998), these data are different from the Colorado Adoption Project (САР), pointing at genetic changes and emergence of new genetic influences at around 7 years of age (Cardon et al., 1992; Fulker et al., 1993). When in other studies shared environmental factors were identified, they appeared to be stable as well (for example, Cardon et al., 1992; Cherny et al., 1998; Fulker et al., 1993). It is assumed that stable family characteristics such as socioeconomic status can impact this effect (Cardon et al., 1992). Our analysis showed that non-shared environment only impact ontogenetic changes in IQ, which is in compliance with literature data.
As we can see sufficiently complex interrelationships were revealed for heritability estimates. We saw the preservation of the same genetic influences during the analyzed age period as well as emergence at the age of 16 of new genetic influences, specific for this age. Probably the appearance of new genetic factors is related to qualitative changes in cognitive development unfolding at this age.
These data showing age dynamics of genetic control of IQ also suggest the dependence of phenotypic variance structure on psychological structure of a studied trait, because the mechanisms of execution of psychological function change in ontogenesis. It becomes especially evident at the beginning of schooling, influencing child’s cognitive activity.
Therefore the accumulating experimental data allow putting forward a question about the dependence of architecture of genetic and environmental influences in individual differences of psychological and psycho-physiological traits from a “context”, in which a trait is realized, its place, meaning, functional role in the whole system, in which it is included at the moment of study. Of course these data only raises new questions and new experimental schemes of behavioral genetic studies using logic of psychological analysis are required.
Thus, according to a number of researchers, new paradigms are needed to explain behavior (Risch & Botstein, 1996, cited in Beckwith & Alper, 2002). An earlier paradigm, which was limited to genetic explanations, didn’t lead to a considerable progress in understanding of the nature of behavior. J. Beckwith and J. Alper suppose that a foundation of a new paradigm should build the idea of complexities caused by genotype-environmental interactions and covariance. Here one should be reminded that genes only act in an environmental context. Because of genotype-environmental covariance and interaction, typical for every complex trait, it seems unlikely that we can precisely estimate the impact of genes on a trait if they are taken away from the environmental context (Beckwith & Alper, 2002).
An interesting fact suggesting genotype-environmental interaction has been reported in a study of Dutch adolescent twins. This study showed that religious education considerably lowers genetic influence on disinhibition, a trait which is related to the “novelty seeking” trait and the drug use (Boomsma, de Geus, van Baal, & Kopmans, 1999).
Unfortunately, oft en enough in sociobiology and behavioral genetics researchers assume that “nature” exists separately from “culture and education”. At the same time many note that “nature” and “culture” are parts of an integrated system of a higher level. Different “developmental sources” reciprocally influence each other. Organism influences its environment and receives a feedback from this environment. A prominent genetic researcher T. Dobzhansky was absolutely correct noting that “Human behavior ... is a result of developmental process, where genes and environment are components of a feedback system” (Dobzhansky, 1967, р. 43).
The idea of circularity shifts a focus of scientists’ attention to characteristics of relationships between the organism and environment, forming the organization of this system. Humans therefore themselves predispose their development (Lerner & Walls, 1999). Besides this, one can’t ignore the idea of intentionality, purposeful and goal-oriented self-development which is getting increasingly influencial on developmental psychology. The basic assumption of this conception is that individuals are at the same time products and creators of their own ontogenetic and personality development in their lifetime. We can’t adequately understand individual ontogenetic development at all life stages without looking at the means by which people (in cognition, activity and social interaction) construct their personal development. Intentionality and intentional world, in which a goal-directed the outside world oriented activity is embedded, become necessary explanatory categories in developmental research (Brandstedter, 1999).
The logic of behavior genetic research should take into account that active individual (the object of investigation) is in relationship with an active context and all events occur in “complexes of contexts” (according to K. Popper). The life path is multidimensional and is not reduced to one trajectory. One of the sources of constant changes is activity conditioning and relating internal, biological, and external, ecological, levels of contexts, in which organism exists.
References
Beckwith, J., & Alper, J.S. (2002). Genetic of Human Personality. Social and Ethical Implications. In J. Benjamin, R. Ebstein, & R. Belmaker (Eds). Molecular Genetics and the human Personality (pp. 315-331). Washington, D.C., London, England: American Psychiatric Publishing.
Brandtstadter, J. (1999). The self in action and development. In Action and Self-Development: Theory and Research through the life span (p. 37-65). Thousand Oaks, London: Sage Publications.
Boomsma, D., van Baal, G.C.M., & Orlebeke J. (1990). Genetic influence on respiratory sinus arrhythmia across different task condions. Acta Geneticae medicae et gemellologiae, 39, 181-191.
Boomsma, D. (1992). Quantitative genetic analyses of cardivascular risk factors in twins and their parents. Amsterdam: Febodruk Enschede.
Boomsma, D.I., & Van Baal, G.C.M. (1998). Genetic influences on childhood IQ in 5- and 7-year-old Dutch twins. Developmental Neuropsychology, 14(1), 115-126.
Boomsma, D., de Geus, E., van Baal, G.C.M., & Kopmans, J. (1999). A religious upbringing reduces the influences of genetic factors on disinhibition: evidence for interaction between genotype and environment on personality. Twin Research, 2, 115-125.
Cardon, L.R., Fulker, D.W., DeFries, J.C., & Plomin, R. (1992). Continuity and change in general cognitive ability from 1 to 7 years of age. Developmental Psychology, 28(1), 64-73.
Cherny, S.S., Fulker, D.W., Emde, R.N., Robinson, J., Corley, R.P., Reznick, J.S., Plomin, R., & DeFries, J.C. (1994). A developmental-genetic analysis of continuity and change in the Bayley Mental Development Index from 14 to 24 months: The MacArthur Longitudinal Twin Study. Psychological Science, 5(6), 354-360.
Dobzhansky, T. (1967). Of flies and men. American Psychologist, 22, 41-48.
Eaves, L., Long, J., & Heath, A. (1986). A theory of developmental quantitative phenotypes applied to cognitive development. Behavior Genetics, 16, 143-162.
Edelman, G.M. (1992). Bright air, brilliant fire. New York: Basic Books.
Egorova, M.S. (2000). Razvitie kak predmet psihogenetiki: rol’ genotipa i sredy v vozrastnyh izmenenijah struktury psikhologicheskih priznakov [Development as a subject of psychogenetics: the role of genotype and environment in the age-related changes of psychological traits]: Avtoref. dis. … d-ra psikhol. nauk. Moscow.
Fulker, D.W., Cherny, S.S., & Cardon, L.R. (1993). Continuity and change in cognitive development. In R. Plomin & G.E. McClearn (Eds.), Nature and Nurture and Psychology (pp. 77-97). Washington, DC: American Psychological Association.
Guttman, L. (1954). A new approach to factor analysis: The radex. In P.F. Lazarsfeld (Ed.). Mathematical thinking in the social sciences (pp. 258-349). Glencoe, Ill.: Free ress.
Ivanchenko, S.N. (1995). Priroda mezhindividual’noj variativnosti parametrov sensomotornoj dejatel’nosti [The nature of interindividual variability of the parameters of the sensorimotor activities]: Avtoref. dis. … kand. psikhol. nauk. Moscow.
Ivanchenko, S.N., & Malykh, S.B. (1994). Priroda izmenchivosti skorostnyh harakteristik sensomotornyh reakcij v razlichnyh jeksperimental’nyh situacijah [Nature of variability in the speed characteristics of sensorimotor reactions in various experimental situations]. Voprosy psikhologii, 6, 80-86.
Kondratieva, I.I. (1973). O nekotoryh osobennostjah projavlenija “zakona sily” v uslovijah specifi cheski chelovecheskoj dejatel’nosti [Some features of manifestations of the “law of force” in a specifi cally human activity]. Voprosy psihologii, 2, 26-36.
Konopkin, O.A. (1980). Psikhologicheskie mehanizmy reguljacii dejatel’nosti [Psychological mechanisms of regulation of activity]. Moscow: Pedagogika.
Kochubei, B.I. (1983). Vlijanie genotipa i sredy na formirovanie individual’nyh osobennostej orientirovochnoj reakcii cheloveka [Influence of genotype and environment on the formation of the individual characteristics of the orienting reaction of human]: Avtoref. dis. … kand. psikhol. nauk. Moscow.
Kochubei, B.I. (1988). Psikhogeneticheskoe izuchenie sistemnyh processov [Psychogenetic study of systemic processes]. In I.V. Ravich-Scherbo (Ed.), Rol’ sredy i nasledstvennosti v formirovanii individual’nosti cheloveka [The role of environment and heredity in shaping human personality] (pp. 138-157). Moscow: Pedagogika.
Kuravsky, L.S., & Malykh, S.B. (2000). On the application of queueing theory for analysis of twin data. Twin Research, 2(3). 92-98.
Kuravsky, L.S., & Malykh, S.B. (2001). Analiz dinamiki sredovyh vlijanij v bliznecovyh issledovanijah [Analysis of the dynamics of environmental infl uences in twin studies]. Voprosy psikhologii, 6, 80-88.
Lerner, R., & Walls, T. (1999). Revisiting individuals as producers of their development. From dynamic interactionism to developmental systems. In Action and Self- Development. Theory and Research through the life span (p. 3-36). Thousand Oaks, London: Sage Publications.
Luria, A.R. (1962). Ob izmerenii psikhologicheskih funkcij v processe razvitija rebenka [On a measure of psychological functions in the process of child development]. Voprosy psikhologii, 3, 15-22.
Luria, A.R. (2002). O prirode psikhologicheskih funkcij i ee izmenchivosti v svete geneticheskogo analiza [About the nature of psychological functions and its variability in the light of genetic analysis]. Voprosy psikhologii, 4, 4-18.
Malykh, S.B. (1986). Individual’nye osobennosti potencialov mozga, svjazannyh s dvizheniem, i rol’ genotipa v ih formirovanii [Individual characteristics of potentials of the brain associated with movement and the role of genotype in their formation]: Avtoref. dis. … kand. psikhol. nauk. Moscow.
Malykh, S.B. (1988). Geneticheskie i sredovye determinanty v izmenchivosti dvigatel’nyh funkcij [Genetic and environmental determinants of variability in motor function]. In I.V. Ravich-Scherbo (Ed.), Rol’ sredy i nasledstvennosti v formirovanii individual’nosti cheloveka [The role of environment and heredity in shaping human personality] (pp. 157-179). Moscow: Pedagogika.
Malykh, S.B. (2000). Geneticheskie osnovy individual’no-psikhologicheskih razlichij: razvitie i struktura psikhologicheskih i psikhofiziologicheskih priznakov [Genetic basis of Individually-psychological differences: the development and structure of the psychological and psychophysiological symptoms]: Avtoref. dis. … d-ra psikhol. nauk. Moscow.
Malykh, S.B., & Kuznecova, I.V. (1992). Struktura fenotipicheskoj dispersii potenciala gotovnosti [The structure of phenotypic variance of the potential availability]. Genetika, 28(11), 160-163.
Malykh, S.B., Kuznecova, I.V., & Sushko, I.N. (1992). Potencialy mozga, svjazannye s dvizheniem u detej 6-7 let [Brain potentials associated with movement for children 6-7 years]. Zhurnal vysshej nervnoj dejatel’nosti, 42(5).
Malykh, S.B., & Orekhova, E.V. (1995). Vozrastnaja dinamika potencialov mozga svjazannyh s dvizheniem u detej 9-12 let [Age dynamics of brain potentials related to motion in children 9-12 years]. Zhurnal vysshej nervnoj dejatel’nosti, 45(2), 269-278.
Malykh, S., & Ravich-Scherbo, I. (1986). Genotypical dependence of the Movement-Related Brain Potentials. In V. Gallai (Ed), Maturation of CNS and Evoked Potentials (p. 247-252). Amsterdam: Elsevier Science Publishers.
Malykh, S.B., Egorova, M.S., & P’yankova, S.D. (1993a). Determinanty individual’nyh osobennostej kognitivnyh harakteristik i psihologicheskaja struktura dejatel’nosti [Determinants of individual features of cognitive performance and psychological structure of activity]. Psikhologicheskij zhurnal, 15(2). 74-84.
Malykh, S.B., Egorova, M.S., & P’yankova, S.D. (1993b). The etiology of individual differences in cognitive strategy. Journal of Russian and East European Psychology, 31 (3), 47-55.
Malykh, S.B., Zyrianova, N.M., & Kuravsky, L.S. (2003). Longitudinal Genetic Analysis of Childhood IQ in 6- and 7-year-old Russian Twins. Twin Research, 6 (4), 285-291.
Maryutina, T.M. (1978). O genotipicheskoj obuslovlennosti vyzvannyh potencialov cheloveka [About genotypic condition evoked potentials of human]. In B.F. Lomov, & I.V. Ravich-Scherbo (Eds.), Problemy geneticheskoj psihofi ziologii cheloveka [Problems of genetic psychophysiology of human] (pp. 72-93). Moscow: Nauka.
Maryutina, T.M. (1988). Priroda mezhindividual’noj izmenchivosti vyzvannyh potencialov [The nature of interindividual variability are evoked potentials]. In I.V. Ravich-Scherbo (Ed.), Rol’ sredy i nasledstvennosti v formirovanii individual’nosti cheloveka [The role of environment and heredity in shaping human personality] (pp. 107- 138). Moscow: Pedagogika.
Maryutina, T.M. (1994). Psikhologicheskie faktory kak determinanty genotip-sredovyh sootnoshenij [Psychological factors as determinants of genotype-environmental relations]. Psikhologicheskij zhurnal, 15(2), 74-84.
Michel G., & Moore, C. (1995). Developmental Psychobiology. An Interdisciplinary Science. (A Bradford Book). Cambridge, Mass., London: The MIT Press.
Olshannikova, A.E. (1962). Projavlenie “zakona sily” v uslovijah, sopostavimyh I nesopostavimyh s uslovijami raboty operatora [Manifestation of the “law of force” in conditions that are comparable and incomparable to the terms of the operator]. Voprosi psikhologii, 5, 31-44.
Orekhova, E.V. (1996). Potencialy mozga, svjazannye s dvizheniem: vozrastnaja dinamika i priroda mezhindividual’noj variativnosti [Potentials of the brain related to motion: Age dynamics and nature of interindividual variability]: Avtoref. dis. … kand. psikhol. nauk. Moscow.
Panteleyeva, T.A. (1977). Analiz individual’nyh razlichij v sensomotornyh reakcijah cheloveka metodom bliznecov [Analysis of individual differences in sensorimotor reactions by method of human twins]: Avtoref. dis. … kand. psikhol. nauk. Moscow.
Panteleyeva, T.A., & Shljahta, N.F. (1978). K issledovaniju genotipicheskoj determinirovannosti nekotoryh pokazatelej labil’nosti nervnyh processov [To study of genotypic determinism of some indicators of lability of nervous processes]. In B.F. Lomov, & I.V. Ravich-Scherbo (Eds.), Problemy geneticheskoj psikhofi ziologii cheloveka [Problems of genetic psychophysiology of human] (pp. 127-135). Moscow: Nauka.
Patrick, C.L. (2000). Genetic and environmental influences on the development of cognitive abilities: Evidence from the field of developmental behavior genetics. Journal of School Psychology, 38(1), 79-108.
Ravich-Scherbo, I.V. (1988). Predislovie [Preface] In I.V. Ravich-Scherbo (Ed.), Rol’ sredy i nasledstvennosti v formirovanii individual’nosti cheloveka [The role of environment and heredity in shaping human personality] (pp. 3-13). Moscow: Pedagogika.
Talyzina, N.F., Krivcova, S.V., & Muhamatulina, E.A. (1991). Priroda individual’nyh razlichij: opyt issledovanija bliznecovym metodom [The nature of individual differences: the experience of method of twin studies]. Moscow: MGU.
Van Baal, G.C.M. (1997). A genetic perspective on the developing brain. (Electrophysiological indices of neural functioning in five to seven year old twins). Amsterdam: Free University.
Wilson, R.S. (1983). The Louisville Twin Study: Developmental Synchronies in behavior. Child Development, 54, 298-316.
Zyryanova, N.M. (1992). Genotip-sredovye sootnoshenija v izmenchivosti pokazatelej kognitivnoj sfery u detej 6-7 let [Genotype-environmental relationship in the variability of cognitive performance in children 6-7 years]: Avtoref. dis. … kand. psikhol. nauk. Moscow.
Notes
[1] This study was supported by grant GC № 02.740.11.0376 from Ministry of Education and Science of the Russian Federation.
To cite this article: Malykh S.B. (2011). Behavior Genetics in Context of Russian Psychology. Psychology in Russia: State of the Art, 4,108-128
The journal content is licensed with CC BY-NC “Attribution-NonCommercial” Creative Commons license.







